
In this post we present selected parts of the very interesting paper titled “Radiocarbon evidence for maritime pioneer colonization at the origins of farming in west Mediterranean Europe“, by João Zilhão.
A sound dating of the first appearance of agro-pastoral economies across Europe is a basic prerequisite to the evaluation of how, why, and when hunter-gatherer adaptive systems eventually disappeared from most of the continent in prehistoric times. The task, however, has not proved easy. Before the advent of accelerator mass spectrometer (AMS) radiocarbon dating, bulk samples of charcoal, bone, or shell had to be used, and in many instances the resultant dates were at odds with archaeological expectations based on stratigraphy and typology.
In west Mediterranean Europe, problems inherent to the nature of the samples have been compounded by the fact that most archaeological evidence comes from caves and rock shelters. Postdepositional disturbance is commonplace in these kinds of sites; hence, when bulk charcoal samples are used, the association between the dated material and the historical events one wants to date cannot be taken for granted before appropriate critical filters are applied to evaluate the association. When scrutinized from a taphonomic perspective, the very early chronology claimed for the inception of production economies in Mediterranean France and Spain must be rejected. The dates (some as early as ≈8000 BP) for the appearance of domesticates in such Spanish sites as Cueva de La Dehesilla and Cova Fosca were obtained from samples collected in disturbed contexts. The dated material contained in variable proportions late Paleolithic, Mesolithic, and Neolithic bone and charcoal, and the Neolithic artifact assemblages found therein corresponded to Epicardial contexts intruded into the underlying late Pleistocene or early Holocene deposits.
A side issue of the Mesolithic-Neolithic transition as seen from these kinds of sites was the local domestication of goats, suggested for Cova Fosca, and the late Mesolithic acquisition of exotic domestic sheep through long distance exchange mechanisms, suggested for Abri Dourgne and Grotte Gazel, among others. This was a definitional issue, involving the identification as domesticates of juvenile ovicaprid bones recovered in high mountain sites that might in fact belong to young chamois or ibex. Such identifications created the impression that in these regions the Neolithic did not spread as a complete integrated package. Instead, local hunter-gatherers would have gone through a process of piecemeal independent invention, or adoption, of different elements. However, once the taxonomic status of bones attributed to sheep reported for those Languedoc sites was reevaluated, the evidence for domesticates in the Mesolithic vanished. This made it clear that the Cardial and related impressed ware cultures of west Mediterranean Europe represent the simultaneous appearance in the archaeological record of the whole package of features involved in the earliest agropastoral practices of the region: animal and plant domesticates, ceramic vessels, polished stone axes, and village dwelling. None of these have ever been found in secure, undisturbed Mesolithic contexts.
Since originally formulated, these conclusions have remained unchallenged. Their verification removed any empirical foundation for models describing the emergence of the Neolithic in west Mediterranean Europe as resulting from gradual eastwest dispersal of economical and technical innovations. Such models dominated the literature for the last quarter of a century. They also are difficult to reconcile with the fact that there is no evidence for long distance exchange or trade networks through which indigenous hunter-gatherer groups could have acquired the Neolithic package (or any of its individual components) and transformed themselves into agro-pastoral societies. Moreover, no sound and testable ecological or social explanations have been offered for why local Mesolithic people would have been willing to undertake such a transformation.
These facts suggest that the spread of farming in west Mediterranean Europe was associated with some sort of demic diffusion process such as the ‘‘wave of advance’’ model put forward by Ammerman and Cavalli-Sforza. According to their analyses, the annual rate of population growth experienced by frontier farmers can hardly have been greater than 1%, and ethnographically observed rates of migratory activity in similar situations never exceed 2,000 km² per generation. This rate carries the implication that the model will not work if observed rates for the spread of farming are greater than 2
km/year. Ammerman and Cavalli-Sforza calculated that the average rate from Greece to the British Isles was close to 1km/year, which is within the model’s expectations. Although regional variations in this overall rate were noted, their causes were thought to lie in environmental specificities (as in the Alpine area) or lack of data (as in the eastern Linear Band Keramik area and Iberia), and it was not conceived that their order of magnitude could be such as to threaten the validity of the model.
As more results were obtained, however, it became clear that in some cases (such as the Linear Bandkeramik and Cardial spreads) these regional rates were more than just ‘‘somewhat faster’’ or ‘‘somewhat slower’’. Moreover, the style of decoration in ceramics from undated sites in Portugal implies contemporaneity with the earliest Cardial sites of eastern Spain and hence an even faster spread. It was suggested, therefore, that the mechanism best explaining the west Mediterranean process was maritime pioneer colonization, a hypothesis first entertained for southern Portugal by Arnaud and for which the occupation of the Pacific islands provides an analogy.

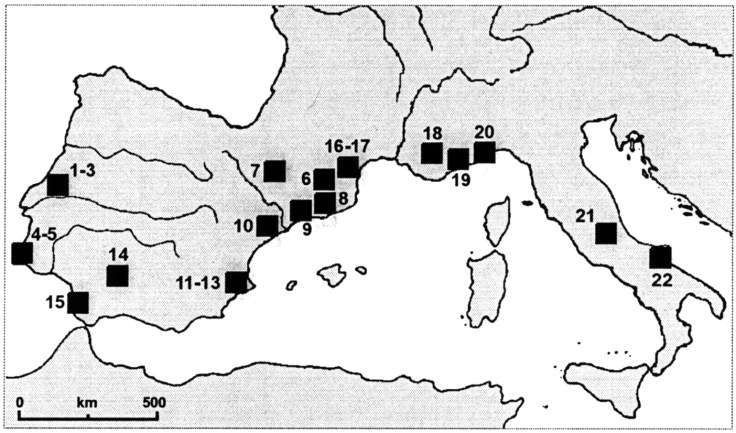
Chronology of the Iberian Cardial Evidence accumulated over the last few years in Iberia shows that eliminating disturbed sites and mixed levels is not sufficient to obtain an adequate evaluation of the rate of spread of the Cardial. There are problems also with the dating of bulk charcoal or wood samples from levels that are unquestionably in situ because of the old wood effect.
Most frequently, this effect is produced by organic material including charcoal, derived from erosion of the surrounding early Holocene soils that contributed to the formation of deposits accumulated in caves or rock shelters in early Neolithic times. In addition, vertically displaced charcoal from underlying Pleistocene levels may penetrate early Neolithic levels even when no signs of disturbance are visible macroscopically and, hence, go unnoticed during excavation. Finally, the use as timber or as fuel of the centuries-old oak trees that constituted the Mediterranean forest cleared by the first agro-pastoral groups may result in the presence in early Neolithic deposits of wood or charcoal material belonging to the inner rings of those trees, the carbon-14 content of which had begun decaying before they were felled or burned. Ages obtained from samples of such material or including such material may be significantly older than the archaeological events for which dating was sought.
A recently published dating experiment demonstrated conclusively that the old wood effect was indeed a major factor to be considered when accounting for the variability in dates from early Neolithic Iberian sites. The first result to be obtained for the Cardial deposits in Cova de les Cendres (Valencia) is 7540±140 BP. The sample was made up of bulk charcoal, and the result was much earlier than the ≈6700 BP that would have been expected based on the dating of the Cardial elsewhere in Spain and France. This result prompted an investigation of the plant species represented in the charcoal assemblage and the consequent identification of Pinus nigra and Juniperus sp., the most common taxa in the underlying Pleistocene levels. Individual pieces of P. nigra and Quercus sp. found in the Cardial deposits were then submitted to AMS dating, with results of 20430±170 BP for the former and 8310±80 BP for the latter.
These results proved beyond a doubt that pieces of Pleistocene and early Holocene charcoal had made their way into the Cardial deposits. Dating a new series of bulk charcoal samples from which P. nigra and Juniperus sp. were excluded provided results closer to what might have been expected and that were accepted as good by the authors of the experiment: 6730±80 BP and 6420±80 BP.
A sample of sheep bone, a short lived diagnostic Neolithic item, was dated to 6280±80 BP. An almost identical result was obtained from charcoal collected in a hearth feature: 6260±80 BP. Both are indistinguishable statistically from a new AMS date of 6340±70 BP for a sample of barley, Hordeum vulgare. Together, these three dates show that the 2σ calibrated age of the earliest Neolithic occupation of Cendres lies in the interval between 5000 and 5500 cal B.C.
Cova de l’Or also represents a fourth demonstration, with Caldeirão, La Draga, and Cendres, that the use of bulk charcoal samples blurs what in fact is a very clear picture. When only results obtained on short lived samples of diagnostic Neolithic items are considered (including those on marine shells collected in Neolithic hearth features excavated at open air sites), all dates for the earliest Cardial, from Valencia to Portugal, are identical statistically. This conclusion receives further support from the new AMS date of 6510±70 BP for wheat seeds from the Cardial levels of the La Falguera rock shelter. Such contemporaneity implies a very rapid spread, within a maximum of four or five generations, of the Iberian Cardial.
Chronology of the Earliest Neolithic in France and Italy The Iberian pattern suggests that radiometric chronology for the earliest Neolithic in neighboring regions to the east also may be skewed significantly by the old wood effect. Unfortunately, this hypothesis is difficult to test, because there are very few direct dates on diagnostic material.
At the open air site of Baratin in Provence, AMS dates on individual pieces of charcoal from three different features provided ages between 6125±80 BP and 6290±70 BP, several centuries younger than the previously available conventional date of 6600±140 BP. At the Arene Candide cave, AMS dating of identified charcoal showed that level 27 yielded both 6150±70 BP remains of Phillyrea sp. and 6880±60 BP remains of Pistacia terebintus. At Coppa Nevigata, two dates on cereal seeds provided virtually identical ages of 6850±80 BP and 6880±90 BP, yet the site was dated previously on conventional bulk charcoal to 7780±320 BP.
The only other site in these regions for which AMS direct dates on diagnostic Neolithic material are available is that of San Marco in Umbria, central Italy, where three samples of Triticum aestivum, Triticum compactum, and H. vulgare seeds provided results of 6430±80 BP, 6270±70 BP, and 6120±90 BP, respectively. Two other conventional results can be considered also as unmistakably related to the early Neolithic use of the sites in question: the 6490±75 BP date on acorns collected inside a burial feature at the Abri Pendimoun and the 6255±255 BP date on human bone from the burial VII of Arene Candide.
When calibrated, these results indicate that the Neolithic begins significantly earlier in southern Italy, perhaps as early as 6000 cal B.C. but that northward and westward all available dates on short lived diagnostic samples are identical to those for Iberia.
Therefore, the rapidity of spread mentioned above for Iberia applies to the dispersal of the Cardial and related cultures as a whole: 2,000 km from the gulf of Genoa to the estuary of the Mondego in probably no more than 100–200 years at most, that is, at a rate of at least 10–20 km/year. It should be stressed that this conclusion holds even if the French chronology based on conventional bulk charcoal samples is retained, because identical results on the same kinds of samples from in situ Cardial deposits are known in Portugal, not only at Caldeirão but also at Pena d’Água, where a sample of Olea charcoal recently has been dated to 6775±60 BP. In any case, these data make it clear that the chronology of the early Neolithic in France and Italy must be revised entirely on the basis of more direct dates of domesticates or on other diagnostic short lived Neolithic materials such as bone artifacts or human bone from burials.

Conclusions By using Ammerman and Cavalli-Sforza’s equations and an annual rate of population growth of 1%, one can calculate a rate of spread of 10 km/year, implying a rate of migratory activity of 60,000 km² per generation, 30 times greater than the maximum observed ethnographically. Demic diffusion therefore cannot have proceeded through a wave of advance mechanism of short distance settlement expansion wherein population growth was accommodated through gradual and slow incorporation of adjacent land.
The area covered by the 40–50-km-wide coastal strip between the Mondego river in Portugal and the cape of Nao, the southern limit of the gulf of Valencia, is precisely 60,000 km². In the framework of maritime pioneer colonization, the similarities in ornaments and pottery decoration observed between the two extremes of this range can be taken as evidence for a swift expansion with maintenance of cultural traditions, and the littoral placement of settlements is evidence for diffusion through sea routes. In fact, under reasonable estimates of annual population growth, the observed rate of spread across such a large area requires the operation of long distance relocation episodes. It also implies such low population densities across the whole of the settled range that large voids must be postulated between nodes of the farmers’ settlement network.
This prediction is met in the archaeological record by the ‘‘enclave’’ nature of early Neolithic territories in littoral-central Portugal, which occupy areas previously uninhabited by late Mesolithic hunter-gatherers. The latter continue to thrive in their own territories for some 500 years after initial Neolithic settlement, but contrasts in material culture, economy, nutrition, and mortuary behavior remain unchanged throughout this period. Neolithic sites feature pottery, polished hand axes, and heat pretreatment of flint, which are absent altogether from the Mesolithic, as are domesticates.
Neolithic skeletons feature isotopic signatures of a fully terrestrial diet, whereas Mesolithic ones indicate a 50% marine component. Neolithic people are buried in special-purpose collective funerary sites, whereas Mesolithic people are buried individually in habitation sites and have never been found carrying exclusively Neolithic body ornaments (tear-shaped Glycymeris beads, pierced red deer canines, and bone beads imitating them).
The rapidity of the spread also indicates that long distance colonization events took place well before saturation levels were attained at the point of origin.

Research-Selection for NovoScriptorium: Philaretus Homerides

Leave a comment