
This post is a presentation of various recent genetic researches that attempt to answer crucial questions about Human Evolution in Europe.

Abstract I review the evolutionary history of human populations in Europe with an emphasis on what has been learned in recent years through the study of ancient DNA. Human populations in Europe ~430-39kya (archaic Europeans) included Neandertals and their ancestors, who were genetically differentiated from other archaic Eurasians (such as the Denisovans of Siberia), as well as modern humans. Modern humans arrived to Europe by ~45kya, and are first genetically attested by ~39kya when they were still mixing with Neandertals. The first Europeans who were recognizably genetically related to modern ones appeared in the genetic record shortly thereafter at ~37kya. At ~15kya a largely homogeneous set of hunter-gatherers became dominant in most of Europe, but with some admixture from Siberian hunter-gatherers in the eastern part of the continent. These hunter-gatherers were joined by migrants from the Near East beginning at ~8kya: Anatolian farmers settled most of mainland Europe, and migrants from the Caucasus reached eastern Europe, forming steppe populations. After ~5kya there was migration from the steppe into mainland Europe and vice versa. Present-day Europeans (ignoring the long-distance migrations of the modern era) are largely the product of this Bronze Age collision of steppe pastoralists with Neolithic farmers.

The oldest sampled nuclear DNA from Europe dates to ~430kya from Sima de los Huesos in Spain and it was found to be more closely related to Neandertals than to Denisovans, unlike mtDNA from the same population, which formed a clade with Denisovan mtDNA. Genomewide data from several European Neandertal individuals has been published down to ~39kya. The Neandertal population that contributed DNA to all non-Africans was more closely related to European Neandertals than to an early (~150kya?) Neandertal from the Altai region in Siberia. Thus, the greater part of the history of European populations was dominated by the Neandertals and their ancestors, raising questions about why this population was replaced in what amounts to a geological blink of an eye.
The genetic divergence between modern humans and Neandertals is lower-bounded by the finding that the Sima de los Huesos hominins already belonged to the Neandertal lineage which must therefore have been already in existence by ~430kya. Modern human and Neandertal Y-chromosomes shared a most recent common ancestor ~450 800kya also pointing to an earlier split of the two lineages. Neandertal mtDNA shared a most recent common ancestor ~270kya, with the earliest known split represented by a specimen from Hohlenstein–Stadel in Germany.
It is only by ~39-36kya that the first sample that clearly shares ancestry with Europeans but not East Asians is evident (Kostenki14 in European Russia), with the earliest known such sample from western Europe at ~35-34kya (GoyetQ116-1 from Belgium). Did these and other early Europeans represent a migration into Europe post the Campanian Ignimbrite (CI) volcanic eruption ~39kya, or are they survivors of this event which set off a short period of intense cooling? The ~42-37kya sample from Oase1 in Romania is the earliest known modern human from Europe, and may predate this event. Oase1 has an excess of 6-9% Neandertal ancestry within a genealogical timeframe of 4-6 generations and no specific affinity to Europeans, suggesting that at least some of the pre-CI Europeans were replaced after this event. It may be that both the modern human and Neandertal inhabitants of Europe suffered a common demise ~39kya.
Intriguingly, GoyetQ116-1 and Kostenki14—two of the earliest samples on the lineage leading to later Europeans—were not symmetrically related to non-Europeans, with GoyetQ116-1 being genetically closer to a ~40kya sample from China (Tianyuan). In a similar vein, mtDNA haplogroup M, rare in Europe today, but common in eastern non-Africans was present in preLast Glacial Maximum Europeans. Surprisingly, this eastern non-African affinity disappeared in samples from the Gravettian-associated “Vestonice-cluster” ~31-26kya which included samples from Italy, Belgium, and the Czech Republic, but partially re-appeared in the ensuing Magdalenian-associated “El Mirón Cluster” ~19-15kya known from sites in Spain, France, Belgium and Germany.
Western European hunter-gatherers (WHG), first described in three sites of western Europe are now known to also have lived in southeastern Europe, Switzerland, the
Baltics, and Italy; the early example from Villabruna in Italy ~15kya has given
this population the alternative name “Villabruna cluster”. The appearance of WHG ~15kya corresponds to the Bølling-Allerød interstadial warm period, and marked a genetic attraction of European and Near Eastern populations. Was this due to migration between Europe and the Near East during this favorable climatic period or due to the expansion of related populations in Europe and the Near East that had been established there at an earlier period? WHG-like ancestry may represent a partial source of ancestry of populations bordering Europe, for example in Anatolia whose early farmers ~8kya are genetically closer to WHG than other Near Eastern populations are, or in the Atlantic where pre-colonial Guanche inhabitants of the Canary Islands had some European hunter-gatherer affinity in addition to their mainly North African origin. WHG did not, however, appear to make any quantifiable genetic contribution to the Upper Paleolithic inhabitants of geographically proximate Morocco ~15kya in North Africa.
Eastern European hunter-gatherers (EHG), a population of mixed WHG and Upper Paleolithic Siberian ancestry (related to the Mal’ta and AfontovaGora specimens from Lake Baikal ~24-17kya) are attested in European Russia ~8kya. This group contributed ancestry to hunter-gatherers in Sweden ~8-5kya, Norway, the Balkans and Ukraine, and
the Baltic. The spread of this ancestry across northern Europe was followed by >3.5kya by the spread of Siberian ancestry that seems to be associated with Finno-Ugrian speakers.
The dominance of the WHG across much of mainland Europe was relatively short-lived, as they were largely replaced beginning in the 7th millennium BC by farmers from Anatolia via southeastern Europe, who minimally, but variably, mixed with incoming farmers in southeastern Europe and propagated their ancestry as far as Scandinavia and Iberia. The WHG populations were, however, persistent, with individuals of
predominantly WHG ancestry found in early Neolithic contexts in Hungary and as late as the 4th millennium BC in the Blätterhöhle site in Germany. These WHG survivors
engendered a resurgence of hunter-gatherer ancestry across Middle Neolithic Europe which appears to have involved local populations of hunter-gatherers rather than a migration from a single WHG-rich area.
Steppe populations during the Eneolithic to Bronze Age were a mix of at least two elements, the EHG who lived in eastern Europe ~8kya and a southern population element related to present-day Armenians, and ancient Caucasus hunter-gatherers, and farmers from Iran. Steppe migrants made a massive impact in Central and Northern Europe post5kya. Some of them expanded eastward, founding the Afanasievo culture and also eventually reached India. These expansions are probable vectors for the spread of Late Proto-Indo-European languages from eastern Europe into both mainland Europe and parts of Asia, but the lack of steppe ancestry in the few known samples from Bronze Age Anatolia raises the possibility that the steppe was not the ultimate origin of Proto-Indo-European (PIE), the common ancestral language of Anatolian speakers, Tocharians, and Late Proto-Indo Europeans.
Our understanding of the spread of steppe ancestry into mainland Europe is becoming
increasingly crisp. Samples from the Bell Beaker complex are heterogeneous, with those
from Iberia lacking steppe ancestry that was omnipresent in those from Central Europe, casting new light on the “pots vs. people” debate in archaeology, which argues that it is dangerous to propose a tight link between material culture and genetic origins. Nonetheless, it is also dangerous to dismiss it completely. Recent studies have shown that people associated with the Corded Ware culture in the Baltics were genetically similar to those from Central Europe and to steppe pastoralists, and the people associated with the Bell Beaker culture in Britain traced ~90% of their ancestry to the continent, being highly similar to Bell Beaker populations there. Bell Beaker-associated individuals were bearers of steppe ancestry into the British Isles that was also present in Bronze Age Ireland, and Iron Age and Anglo-Saxon England. The high genetic similarity between people from the British Isles and those of the continent makes it more difficult to trace migrations into the Isles. This high similarity masks a very detailed fine-scale population structure that has been revealed by study of present-day individuals; a similar type of analysis applied to ancient DNA has the potential to reveal fine-grained population structure in ancient European populations as well.
Southeastern Europe received steppe-related ancestry before any other population in Europe outside the steppe itself, with sporadic appearance of individuals with steppe ancestry in Bulgaria as early as ~6.7-6.5kya and a general low-level presence of ~30% during the Bronze Age, ~5.4-3.1kya. This ancestry was also present in the Aegean during the Mycenaean period ~3.5kya at ~15%, but was absent from the otherwise genetically similar Minoan culture of Crete who represents the most recent sampled European population without any such ancestry. Both Minoans and Mycenaeans, and to a much lesser extent Neolithic samples from the Peloponnese and Bulgaria also had ancestry related to Caucasus hunter-gatherers, suggesting that this ancestry did not come to Europe only via migrations from the steppe, but also independently, perhaps reflecting ancestry from different Anatolian source populations.

Ancient DNA research has helped us reconstruct many previously unsuspected events in
European evolutionary history, but it also opened up many new questions: Where did the first European-like humans (represented by Kostenki14 and GoyetQ116-1) arrive from? What contributed to the genetic differentiation between Europe and the Near East; how did Near Eastern populations acquire Basal Eurasian ancestry and why did this ancestry not spread into geographically neighboring Europe? How did the WHG become so successful in Europe ~15kya virtually replacing all previous Ice Age Europeans? What drove ancient Siberian populations westward to produce the European/Siberian intermediate EHG? What was the mechanism by which steppe populations of mixed EHG and Near Eastern ancestry were formed, and what was the mechanism by which steppe populations managed to make a major demographic impact in Neolithic Europe?
(Source: “The evolutionary history of human populations in Europe”, by Iosif Lazaridis)

Abstract Farming and sedentism first appear in southwest Asia during the early Holocene and later spread to neighboring regions, including Europe, along multiple dispersal routes. Conspicuous uncertainties remain about the relative roles of migration, cultural diffusion and admixture with local foragers in the early Neolithisation of Europe. Here we present paleogenomic data for five Neolithic individuals from northwestern Turkey and northern Greece, spanning the time and region of the earliest spread of farming into Europe. We observe striking genetic similarity both among Aegean early farmers and with those from across Europe. Our study demonstrates a direct genetic link between Mediterranean and Central European early farmers and those of Greece and Anatolia, extending the European Neolithic migratory chain all the way back to southwestern Asia.

It is well established that farming was introduced to Europe from Anatolia, but the extent to which its spread was mediated by demic expansion of Anatolian farmers, or by the transmission of farming technologies and lifeways to indigenous hunter-gatherers without a major concomitant migration of people, has been the subject of considerable debate. Paleogenetic studies of late hunter-gatherers (HG) and early farmers indicate a dominant role for migration in the transition to farming in central and northern Europe, with evidence of only limited hunter-gatherer admixture into early Neolithic populations, but increasing toward the late Neolithic. However, the exact origin of central and western Europe’s early farmers in the Balkans, Greece, or Anatolia remains an open question.
Recent radiocarbon dating indicates that by 6,600–6,500 calibrated (cal) BCE sedentary farming communities were established in northwestern Anatolia at sites such as Barcın, Menteşe, and Aktopraklık C and in coastal western Anatolia at sites such as Çukuriçi and Ulucak, but did not expand north or west of the Aegean for another several hundred years. All these sites show material culture affinities with the central and southwestern Anatolian Neolithic.
Early Greek Neolithic sites, such as the Franchthi Cave in the Peloponnese, Knossos in Crete, and Mauropigi, Paliambela, and Revenia in northern Greece date to a similar period. The distribution of obsidian from the Cycladic islands, as well as similarities in material culture, suggest extensive interactions since the Mesolithic and a coeval Neolithic on both sides of the Aegean. Although it has been argued that in situ Aegean Mesolithic hunter-gatherers played a major role in the “Neolithization” of Greece, the presence of domesticated forms of plants and animals indicates nonlocal Neolithic dispersals into the area.
It is widely believed that farming spread into Europe along both Mediterranean and central European routes, but the extent to which this process involved multiple dispersals from the Aegean has long been a matter of debate.
Ötzi the Tyrolean Iceman shows unique shared drift with Aegeans to the exclusion of Hungarian Early Neolithic farmers and Late and Post Neolithic European genomes and feasibly represents a relict of Early Neolithic Aegeans.
Given that the Aegean is the likely origin of European Neolithic farmers, we used Bar8 and Bar31 as putative sources to assess the extent of hunter-gatherer admixture in European farmers through the Neolithic. f4 statistics of the form f4 (Neolithic farmer, Anatolian, HG, ‡Khomani) indicated small but significant amounts of hunter-gatherer admixture into both Spanish and Hungarian early farmer genomes, and interestingly, the Early Neolithic Greek genome. Our mixture modeling analysis also inferred a small genetic contribution from the Loschbour hunter-gatherer genome (3–9%) to each of the Early Neolithic Hungarian and German genomes, but evidence of a smaller contribution to any Aegean genomes (0–6%). These results suggest that mixing between migrating farmers and local hunter-gatherers occurred sporadically at low levels throughout the continent even in the earliest stages of the Neolithic. However, consistent with previous findings, both f4 statistics and ADMIXTURE analysis indicate a substantial increase in hunter-gatherer ancestry transitioning into the Middle Neolithic across Europe, whereas Late Neolithic farmers also demonstrate a considerable input of ancestry from steppe populations.
Most of the modern Anatolian and Aegean populations do not appear to be the direct descendants of Neolithic peoples from the same region. Indeed, our mixture model comparison of the Aegean genomes to >200 modern groups indicates low affinity between the two Anatolian Neolithic genomes and six of eight modern Turkish samples; the other two were sampled near the Aegean Sea at a location close to the site of the Neolithic genomes. Furthermore, when we form each Anatolian Neolithic genome as a mixture of all modern groups, we infer no contributions from groups in southeastern Anatolia and the Levant, where the earliest Neolithic sites are found. Similarly, comparison of allele sharing between ancient and modern genomes to those expected under population continuity indicates Neolithic-to-modern discontinuity in Greece and western Anatolia, unless ancestral populations were unrealistically small. Instead, our mixing analysis shows that each Aegean Neolithic genome closely corresponds to modern Mediterraneans (>68% contributions from southern Europe) and in particular to Sardinians (>25%), as also seen in the PCA and outgroup f3 statistics with few substantial contributions from elsewhere. Modern groups matching the Neolithics—mostly from the Mediterranean and North Africa—strikingly match more to Bar8 from northwestern Anatolia than to the LBK genome from Stuttgart in Germany, indicating that the LBK genome experienced processes such as drift and admixture that were independent from the Mediterranean expansion route, consistent with the dual expansion model.

Concluding Remarks The high levels of shared drift between Aegean and all available Early Neolithic genomes in Europe, together with the inferred unique drift between Neolithic Aegeans and Early Neolithic genomes from Northern Spain to the exclusion of Early Neolithic genomes from central Europe, indicate that Aegean Neolithic populations can be considered the root for all early European farmers and that at least two independent colonization routes were followed.
(Source: “Early farmers from across Europe directly descended from Neolithic Aegeans”, by Zuzana Hofmanová et al.)

Abstract Farming was first introduced to southeastern Europe in the mid-7th millennium BCE brought by migrants from Anatolia who settled in the region before spreading throughout Europe. However, the dynamics of the interaction between the first farmers and the indigenous hunter-gatherers remain poorly understood because of the near absence of ancient DNA from the region. We report new genome-wide ancient DNA data from 204 individuals–65 Paleolithic and Mesolithic, 93 Neolithic, and 46 Copper, Bronze and Iron Age–who lived in southeastern Europe and surrounding regions between about 12,000 and 500 BCE. We document that the hunter-gatherer populations of southeastern Europe, the Baltic, and the North Pontic Steppe were distinctive from those of western Europe, with a West-East cline of ancestry. We show that the people who brought farming to Europe were not part of a single population, as early farmers from southern Greece are not descended from the Neolithic population of northwestern Anatolia that was ancestral to all other European farmers. The ancestors of the first farmers of northern and western Europe passed through southeastern Europe with limited admixture with local hunter-gatherers, but we show that some groups that remained in the region mixed extensively with local hunter-gatherers, with relatively sex-balanced admixture compared to the male-biased hunter-gatherer admixture that we show prevailed later in the North and West. After the spread of farming, southeastern Europe continued to be a nexus between East and West, with intermittent steppe ancestry, including in individuals from the Varna I cemetery and associated with the Cucuteni-Trypillian archaeological complex, up to 2,000 years before the Steppe migration that replaced much of northern Europe’s population.
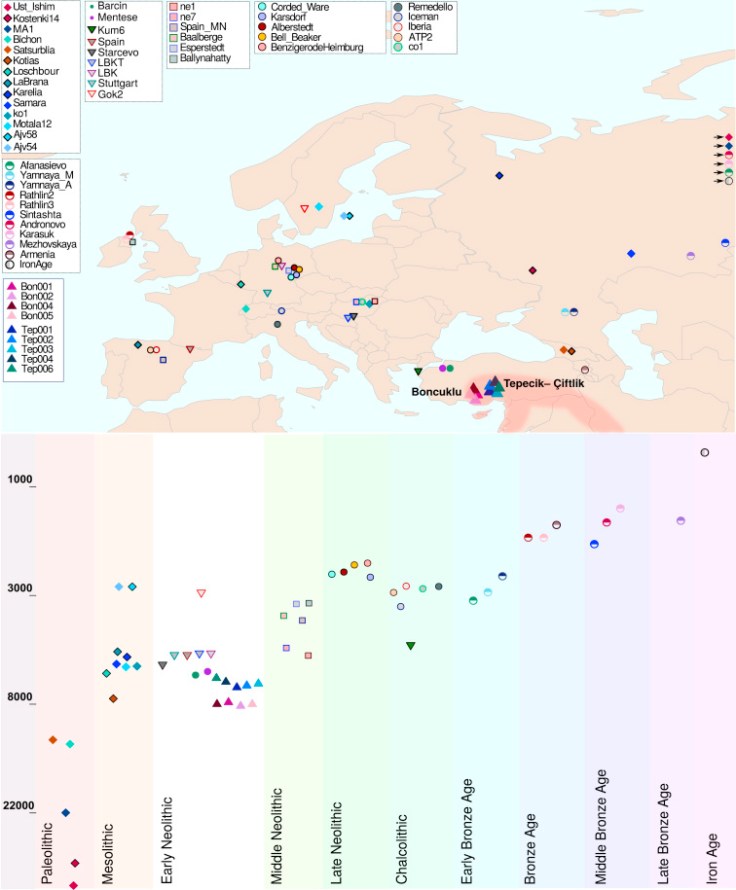
We report new data from hunter-gatherers from France, Sicily and Croatia, as well as higher coverage data from three previously published hunter-gatherers from France and Germany. The Sicilian and Croatian individuals dating to 12,000 and 6100 BCE
cluster closely with western hunter-gatherers, including individuals from Loschbour (Luxembourg, 6100 BCE), Bichon (Switzerland, 11,700 BCE), and Villabruna (Italy 12,000
BCE). These results demonstrate that the “western hunter-gatherer” population was widely distributed from the Atlantic seaboard of Europe in the West, to Sicily in the South, to the Balkan Peninsula in the Southeast, for at least six thousand years, strengthening the evidence that the western hunter-gatherers represent a population that expanded from a southeastern European refugium following the last Ice Age around 15,000 years ago–in the process displacing or admixing with the existing population of western Europe.
Modeling Balkan Neolithic populations as a mixture of Anatolia Neolithic, western hunter-gatherer and Ukraine Mesolithic, we estimate that Balkan Neolithic populations derive 98% (95% 292 confidence interval [CI]; 97-100%) of their ancestry from populations related to those of the northwestern Anatolian Neolithic. A striking exception to the pattern of limited hunter-gatherer admixture in Balkan Neolithic populations is evident in 8 out of 9 individuals from Malak Preslavets, a site in present-day Bulgaria close to the Danube river. These individuals likely lived in the mid-6th millennium BCE and have significantly more hunter-gatherer ancestry than other Balkan Neolithic individuals as shown by PCA and ADMIXTURE as well as D-statistics and qpAdm modeling.
In the Balkans, Copper Age populations have significantly more hunter-gatherer ancestry than Neolithic populations.
We also report the first data from the Late Neolithic Globular Amphora Complex. Globular Amphora individuals from two sites in Poland and Ukraine form a tight genetic cluster, showing genetic homogeneity over a large distance. We find that this population had more hunter-gatherer ancestry than Middle Neolithic groups from Central Europe.
The Globular Amphora abutted populations with steppe-influenced material cultures for hundreds of years and yet the individuals in our study have no evidence of steppe ancestry, suggesting that this persistent culture frontier corresponded to a genetic barrier.
The migrations from the Pontic-Caspian steppe associated with the Yamnaya Cultural Complex in the 3rd millennium BCE made a profound contribution to the genetic ancestry of central Europe, contributing about 75% of the ancestry of individuals associated with the Corded Ware Complex and about 50% of the ancestry of succeeding material cultures such as the Bell Beaker Complex.
We confirm that Mediterranean populations, represented in our study by individuals of the Impressa complex from Croatia and the Epicardial Early Neolithic from Spain, are closely related to the Danubian population represented by the Linearbandkeramik (LBK) from central Europe and show that both groups are closely related to the Balkan Neolithic population. These three populations form a clade with Northwest Anatolians as an outgroup, consistent with a single migration from a population closely related to the northwestern Anatolian Neolithic farming population into the Balkan peninsula, which then split into two populations that followed the Danubian and Mediterranean routes.
A related question about the spread of farming into Europe concerns whether its initial arrival in present-day Greece and subsequent expansion was mediated by a single population migrating from Anatolia – as has been consistent with genetic data up until now – or whether there were multiple initial groups, as suggested by the archaeological record. We find that four southern Greek (Peloponnese) Neolithic individuals – three from Diros Cave and one from Franchthi Cave, plus one previously published individual from Diros – are not consistent with descending from the same source population as other European farmers. In PCA these individuals are outliers; shifted away from northwestern Anatolian and European Early Neolithic individuals, in a direction opposite from WHG. D-statistics show that in fact, these “Peloponnese Neolithic” individuals have less WHG-related ancestry than Anatolia Neolithic ones, and that they form an outgroup relative to Anatolian and Balkans Neolithic populations, suggesting an independent migration into Europe from a population that split off from the ancestors of the northwest Anatolian individuals from which we have data.
Admixture graph modeling supports this interpretation, confirming that their Near Eastern ancestry is derived from a lineage that is close, or basal, to the non-WHG component of Anatolian Neolithic ancestry. One possibility is that this independent migration is related to an earlier Aceramic Neolithic in Greece that was derived from the pre-pottery Neolithic (PPNB) of Cyprus and the Levant. Under this model, the earliest Neolithic populations in Greece migrated from the Levant, perhaps via the southwestern Anatolian coast as early as 7000 BCE, but the majority of Neolithic ancestry arrived around 500 years later via a route that passed through northwestern Anatolia. The predictions of this hypothesis could be further tested with genome-wide data of Early Neolithic individuals from Cyprus, Crete and southwest Anatolia. Populations related to the Peloponnese Neolithic potentially made a small contribution to the ancestry of other Mediterranean Neolithic populations like Early Neolithic Iberia and Neolithic farmers from northern Greece but we do not strongly reject models without such a contribution.

Discussion Our study shows that southeastern Europe consistently served as a genetic contact zone between different populations. This role likely contributed to the extraordinary series of cultural innovations that characterize the region, from the elegant figurines of the Neolithic to the ornaments and precious metalwork of Varna. Before the arrival of farming, this region saw constant interaction between highly diverged groups of hunter-gatherers, and this interaction continued, perhaps accelerating, after the arrival of farming. We find evidence that some early farmers from Greece derived ancestry from a different source compared to the one that contributed the majority of ancestry of all other farmers in Europe. In eastern Europe we document the appearance of CHG/Iranian Neolithic ancestry north of the Black Sea, and its eventual extension as far north as the Baltic. In some ways, this expansion parallels the expansion of Anatolian farmer ancestry into western Europe although it is less dramatic, and several thousand years later. These expansions set up the two, largely separate, populations in western and eastern Europe that would come together in the Final Neolithic and Early Bronze Age to form the ancestry of present-day Europe.
This study describes key ancestral components that contributed to present-day West Eurasian genetic diversity. However, the more recent processes that created present-day southeastern European populations are unknown, and understanding this will require dense sampling of Bronze Age, Iron Age, Roman, and Medieval groups and comparison to present-day populations. At the most ancient end of our time series, while information about hunter-gatherer population structure in northern and western Europe now extends back throughout the Upper Paleolithic, we have little data about how these populations fit into a wider Eurasian context, and more data from hunter-gatherer populations in Anatolia, the Near East and East Asia will be needed to resolve that question. Finally, many questions about the nature of the interactions between populations remain unresolved. For example, we report evidence for sex-bias in one particular set of interactions between hunter-gatherers and farmers, and other interactions may have had similar dynamics. However, many more examples of such interactions need to be collected before it will become possible to make generalizable claims about the patterns of sex-biased interactions among human populations as they came into contact and mixed during prehistory.
(Source: “The Genomic History of Southeastern Europe”, by Mathieson & Reich et al.)

The archaeological documentation of the development of sedentary farming societies in Anatolia is not yet mirrored by a genetic understanding of the human populations involved, in contrast to the spread of farming in Europe. Sedentary farming communities emerged in parts of the Fertile Crescent during the tenth millennium and early ninth millennium calibrated (cal) BC and had appeared in central Anatolia by 8300 cal BC. Farming spread into west Anatolia by the early seventh millennium cal BC and quasi-synchronously into Europe, although the timing and process of this movement remain unclear. Using genome sequence data that we generated from nine central Anatolian Neolithic individuals, we studied the transition period from early Aceramic (Pre-Pottery) to the later Pottery Neolithic, when farming expanded west of the Fertile Crescent. We find that genetic diversity in the earliest farmers was conspicuously low, on a par with European foraging groups. With the advent of the Pottery Neolithic, genetic variation within societies reached levels later found in early European farmers. Our results confirm that the earliest Neolithic central Anatolians belonged to the same gene pool as the first Neolithic migrants spreading into Europe. Further, genetic affinities between later Anatolian farmers and fourth to third millennium BC Chalcolithic south Europeans suggest an additional wave of Anatolian migrants, after the initial Neolithic spread but before the Yamnaya-related migrations. We propose that the earliest farming societies demographically resembled foragers and that only after regional gene flow and rising heterogeneity did the farming population expansions into Europe occur.
(Source: “The Demographic Development of the First Farmers in Anatolia”, by Gülsah Merve Kilinc et al.)
We report genome-wide ancient DNA from 44 ancient Near Easterners ranging in time
between ~12,000-1,400 BCE, from Natufian hunter-gatherers to Bronze Age farmers. We show that the earliest populations of the Near East derived around half their ancestry from a ‘Basal Eurasian’ lineage that had little if any Neanderthal admixture and that separated from other non-African lineages prior to their separation from each other. The first farmers of the southern Levant (Israel and Jordan) and Zagros Mountains (Iran) were strongly genetically differentiated, and each descended from local hunter-gatherers. By the time of the Bronze Age, these two populations and Anatolian-related farmers had mixed with each other and with the hunter-gatherers of Europe to drastically reduce genetic differentiation. The impact of the Near Eastern farmers extended beyond the Near East: farmers related to those of Anatolia spread westward into Europe; farmers related to those of the Levant spread southward into East Africa; farmers related to those from Iran spread northward into the Eurasian steppe; and people related to both the early farmers of Iran and to the pastoralists of the Eurasian steppe spread eastward into South Asia.
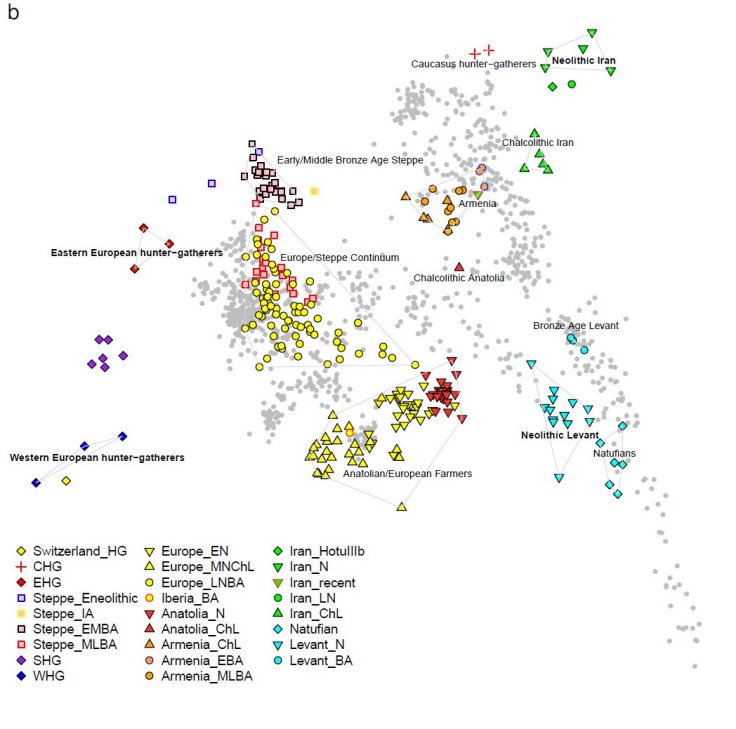
Our data document continuity across the hunter-gatherer / farming transition, separately in the southern Levant and in the southern Caucasus-Iran highlands.
Almost all ancient and present-day West Eurasians have evidence of significant admixture between two or more ancestral populations.
Our analysis show that the ancient populations of the Chalcolithic Iran, Chalcolithic Armenia, Bronze Age Armenia and Chalcolithic Anatolia were all composed of the same ancestral components, albeit in slightly different proportions.
Admixture did not only occur within the Near East but extended towards Europe. To the
north, a population related to people of the Iran Chalcolithic contributed ~43% of the
ancestry of early Bronze Age populations of the steppe.
The early farmers of mainland Europe were descended from a population related to Neolithic northwestern Anatolians.
This is consistent with an Anatolian origin of farming in Europe, but does not reject other sources, since the spatial distribution of the Anatolian/European-like farmer populations is unknown. We can rule out the hypothesis that European farmers stem directly from a population related to the ancient farmers of the southern Levant.
Our analysis shows that East African ancestry is significantly better modelled by Levantine early farmers than by Anatolian or early European farmers, implying that the spread of this ancestry to East Africa was not from the same group that spread Near Eastern ancestry into Europe.
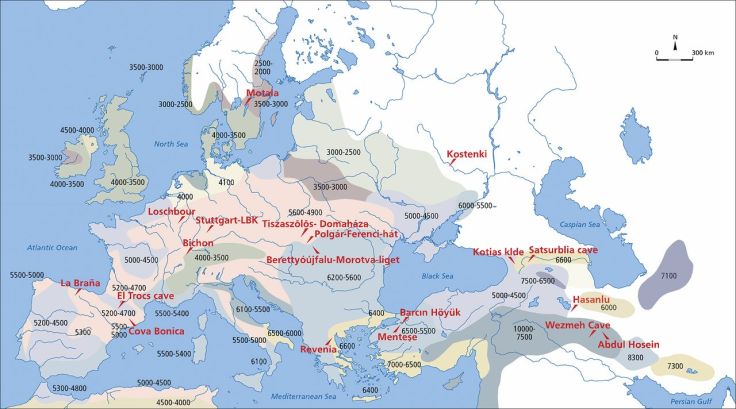
Conclusions By analysing genome-wide ancient DNA data from ancient individuals from the Levant, Anatolia, the southern Caucasus and Iran, we have provided a first glimpse of the demographic structure of the human populations that transitioned to farming. We reject the hypothesis that the spread of agriculture in the Near East was achieved by the dispersal of a single farming population displacing the hunter-gatherers they encountered. Instead, the spread of ideas and farming technology moved faster than the spread of people, as we can determine from the fact that the population structure of the Near East was maintained throughout the transition to agriculture. A priority for future ancient DNA studies should be to obtain data from older periods, which would reveal the deeper origins of the population structure in the Near East. It will also be important to obtain data from the ancient civilizations of the Near East to bridge the gap between the region’s prehistoric inhabitants and those of the present.
(Source: “The genetic structure of the world’s first farmers”, by Iosif Lazaridis et al.)

Abstract In the early seventh millennium BC an expansion of the Neolithic economyand sedentism took place in Asia Minor. This occurred nearly two millennia after theemergence of Neolithic societies in southern central Anatolia, which raises the question of how this expansion occurred, and why it came about at this particular moment. This paperconsiders various elements that might have played a role in this expansion episode, such asclimate change, demography, and agricultural and social changes.
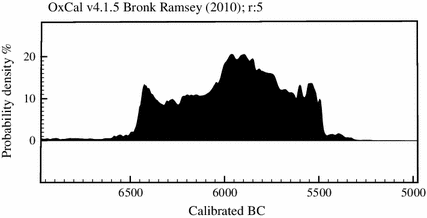
In conclusion, I have argued that the Neolithic expansion in Asia Minor that occurredaround 6500 BC was a creative episode, involving the adoption of new farming technol-ogies by groups that consisted of both migrant farmers and local hunter-gatherers. It isclear then that the idea that Anatolia was simply a bridge transmitting the Neolithic fromthe Fertile Crescent to Europe should be discarded. Finally, I have only scratched thesurface here of the processes occurring in the seventh millennium BC in Asia Minor, whichdeserve much more detailed investigation.
NovoScriptorium: We firmly suggest a read of this whole paper. It is very well written indeed, and at the same time it represents the real situation of our knowledge on human migration; endless, as rational as they may be, assumptions.
(Source: “Breaking the bond: Investigating the Neolithic expansion in Asia Minor in the seventh millennium BC”, by Bleda S. Düring)

Abstract Ancient DNA studies have established that European Neolithic populations were descended from Anatolian migrants who received a limited amount of admixture from resident hunter-gatherers. Many open questions remain, however, about the spatial and temporal dynamics of population interactions and admixture during the Neolithic period. Using the highest-resolution genome-wide ancient DNA data set assembled to date—a total of 177 samples, 127 newly reported here, from the Neolithic and Chalcolithic of Hungary (6000–2900 BCE, n = 98), Germany (5500–3000 BCE, n = 42), and Spain (5500–2200 BCE, n = 37)—we investigate the population dynamics of Neolithization across Europe. We find that genetic diversity was shaped predominantly by local processes, with varied sources and proportions of hunter-gatherer ancestry among the three regions and through time. Admixture between groups with different ancestry profiles was pervasive and resulted in observable population transformation across almost all cultural transitions. Our results shed new light on the ways that gene flow reshaped European populations throughout the Neolithic period and demonstrate the potential of time-series-based sampling and modeling approaches to elucidate multiple dimensions of historical population interactions.

The population dynamics of the Neolithization process are of great importance for understanding European prehistory. The first quantitative model of the Neolithic transition to integrate archaeological and genetic data was the demic diffusion hypothesis, which posited that growing population densities among Near Eastern farmers led to a range expansion that spread agriculture to Europe. Ancient DNA analysis has validated major migrations from populations related to Neolithic Anatolians as driving the arrival of farming in Europe, but the demic diffusion model does not account for the complex-ities of the interactions between farmers and hunter-gatherers in Europe throughout the Neolithic.
Genetic data have yet to be used systematically to model the population interactions and transformations during the course of the Neolithic period. Key open questions include whether migrating farmers mixed with hunter-gatherers at each stage of the expansion, and if so how soon after arriving. Additionally, while it has previously been shown that hunter-gatherer ancestry among farmers in several parts of Europe had increased by the Middle Neolithic, it is currently unclear whether this was a continuous or discrete process and furthermore whether it involved a continent-wide phenomenon or a variety of parallel, local events.
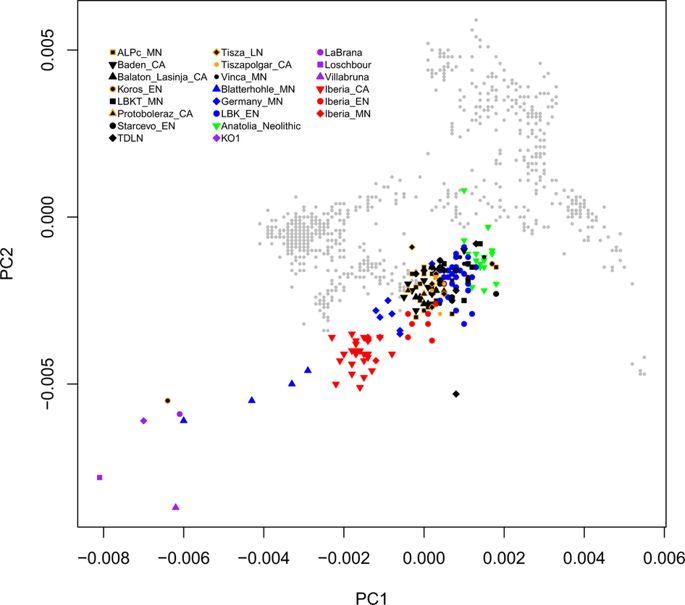
Our results provide greatly increased detail in understanding population interactions and admixture during the European Neolithic. In each of our three study regions (Spain, Germany, Hungary), the arrival of farmers prompted admixture with local hunter-gatherers which unfolded over thousands of years: almost all sampled populations from subsequent time periods have more hunter-gatherer ancestry and more recent dates of admixture than their local predecessors, suggesting recurrent changes in genetic composition and significant hunter-gatherer gene flow beyond initial contact. These transformations left distinct signatures in each region, implying that they likely resulted from a complex web of local interactions rather than a uniform demographic phenomenon. Our transect of Hungary, in particular, with representative samples from many archaeological cultures across the region and throughout the Neolithic and Chalcolithic, illustrates the power of dense ancient DNA time series. Future work with similar data sets and statistical models promises to teach us much more about population transformations in space and time.
(Source: “Parallel ancient genomic transects reveal complex population history of early European farmers”, by Mark Lipson et al.)

Abstract While the series of events that shaped the transition between foraging societies and food producers are well described for Central and Southern Europe, genetic evidence from Northern Europe surrounding the Baltic Sea is still sparse. Here, we report genome-wide DNA data from 38 ancient North Europeans ranging from ~9500 to 2200 years before present. Our analysis provides genetic evidence that hunter-gatherers settled Scandinavia via tworoutes. We reveal that the first Scandinavian farmers derive their ancestry from Anatolia 1000 years earlier than previously demonstrated. The range of Mesolithic Western hunter-gatherers extended to the east of the Baltic Sea, where these populations persisted without gene-flow from Central European farmers during the Early and Middle Neolithic. The arrival of steppe pastoralists in the Late Neolithic introduced a major shift in economy and mediated the spread of a new ancestry associated with the Corded Ware Complex in Northern Europe.
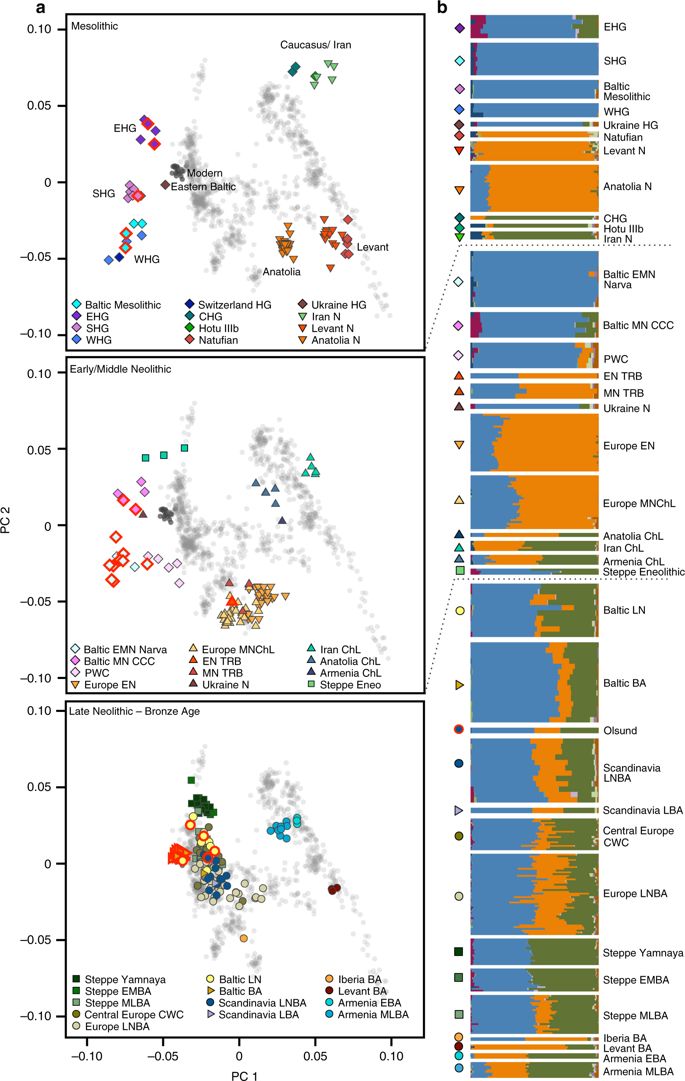
Recent studies of ancient human genomes have revealed acomplex population history of modern Europeans involving at least three major prehistoric migrations, influenced by climatic conditions, the availability of resources, the spread of technological and cultural innovations, and possibly diseases. However, the archaeological record of the very north of the European subcontinent surrounding today’s Baltic Sea shows a history distinct to that of Central and Southern Europe which has not yet been comprehensively studied on a genomic level.
The skeletal remains studied here were recovered from 25 archaeological sites in the territory of modern Lithuania, Latvia, Estonia, Archangelsk Oblast and Karelia (north-western Russia) and Sweden dating from around 7500 to 200 calBCE.
Similarly to the Baltic Mesolithic, the later Eastern Baltic Neolithic hunter-gatherers of the Narva culture exhibit varying proportions of EHG (0– 46%) and WHG (54– 100%) ancestry.
In contrast to the Eastern Baltic, we see clear evidence for the genetic impact of the Neolithic expansion already around 4000 calBCE in southern Sweden.
The substantial population movement at the beginning of the 3rd millennium calBCE, during the European LNBA, affected the genetic make up of Eastern and Central Europe and Scandinavia. It also made its mark in the Eastern Baltic region.
Discussion Our analyses support a dynamic population history of the Baltic Sea region, where populations did not remain in ‘genetic stasis’ despite the late adoption of agrarian subsistence strategies when compared to the rest of Europe.
In southern Scandinavia, the sequence of events resembles that seen in Central Europe, albeit several millennia later, in that the earliest agriculture in the region coincides with the appearance of people related to the Anatolian and European Neolithic. However, similar to Middle Neolithic Central and Southern European populations, early Scandinavian farmers are already strongly admixed with hunter-gatherer groups.
In the archaeological understanding, the transition from Mesolithic to Neolithic in the Eastern Baltic region does not coincide with a large-scale population turnover and a stark shift in economy as seen in Central and Southern Europe. Rather, it is signified by a change in networks of contacts and the use of pottery, among other material, cultural and economic changes.
We see a further population movement into the regions surrounding the Baltic Sea with the CWC in the Late Neolithic that was accompanied by the first evidence of extensive animal husbandry in the Eastern Baltic. The presence of ancestry from the Pontic-Caspian Steppe among Baltic CWC individuals without the genetic component from north-western Anatolian Neolithic farmers must be due to a direct migration of steppe pastoralists that did not pick up this ancestry in Central Europe.
Local foraging societies were, however, not completely replaced and contributed a substantial proportion to the ancestry of Eastern Baltic individuals of the latest LN and Bronze Age.
(Source: “The genetic prehistory of the Baltic Sea region”, by Alissa Mittnik et al.)

Abstract Recent ancient DNA studies have revealed that the genetic history of modern Europeans was shaped by a series of migration and admixture events between deeply diverged groups. While these events are well described in Central and Southern Europe, genetic evidence from Northern Europe surrounding the Baltic Sea is still sparse. Here we report genome-wide DNA data from 24 ancient North Europeans ranging from ~7,500 to 200 calBCE spanning the transition from a hunter-gatherer to an agricultural lifestyle, as well as the adoption of bronze metallurgy. We show that Scandinavia was settled after the retreat of the glacial ice sheets from a southern and a northern route, and that the first Scandinavian Neolithic farmers derive their ancestry from Anatolia 1000 years earlier than previously demonstrated. The range of Western European Mesolithic hunter-gatherers extended to the east of the Baltic Sea, where these populations persisted without gene-flow from Central European farmers until around 2,900 calBCE when the arrival of steppe pastoralists introduced a major shift in economy and established wide-reaching networks of contact within the Corded Ware Complex.
Recent genetic studies of ancient human genomes have revealed a complex population history of modern Europeans involving at least three major prehistoric migrations that were influenced by climatic conditions and availability of resources as well as the spread of technological and cultural innovations and possibly diseases. However, how and when they affected the populations of the very north of the European continent surrounding today’s Baltic Sea, where the archeological record shows a distinct history to that of Central and Southern Europe, has yet to be comprehensively studied on a genomic level.
The archeological record of the eastern Baltic and Scandinavia shows that settlement by mobile foragers started only after retreat of the glacial ice sheets around 11,000 years before present. To the west and south, hunter-gatherers (Western Hunter-Gatherers or WHG) sharing a common genetic signature already occupied wide ranges of Europe from Iberia to Hungary for several millennia. They were shown to be descended from foragers appearing in Europe after around 14,000 years ago in a population turnover coinciding with the warming period of the Bølling-Allerød interstadial, possibly emerging from a southern refugium they inhabited since the Glacial Maximum and replacing preceding foraging populations.
The following Early Neolithic period, starting around 6,000 calibrated radiocarbon years before Common Era (calBCE), saw the transition from foraging to a sedentary agricultural lifestyle with the expansion of farmers out of Anatolia following the Danube and Mediterranean coast into Central and Southern Europe where they existed in parallel and admixed with local foragers for the following two millennia. This development reached South Scandinavia at around 4,000 calBCE with the farmers of the Funnel Beaker Culture (TRB; from German Trichterbecher) who gradually introduced cultivation of cereals and cattle rearing. At the transition to the northern Middle Neolithic, around 3,300 calBCE, an intensification of agriculture is seen in Denmark and western Central Sweden accompanied by the erection of megaliths and changes in pottery and lithic technology, while settlements in eastern Central Sweden increasingly concentrated along the coast and economy shifted toward marine resources such as fish and seal.
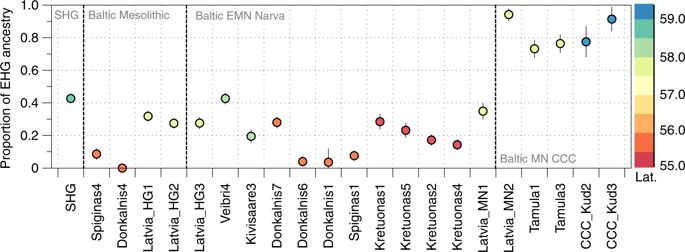
The production and use of pottery, in Central and southern Europe often seen as part of the ‘Neolithic package’, was already common among foragers in Scandinavia during the preceding Mesolithic Ertebølle phase. Similarly in the eastern Baltic, where foraging continued to be the main form of subsistence until at least 4,000 calBCE, ceramics technology was adopted before agriculture. Recent genome wide data of hunter-gatherers from the Baltic Narva Culture revealed genetic continuity with the preceding Mesolithic inhabitants of the same region as well as influence from the more northern EHG.
The Late Neolithic is seen as a major transformative period in European prehistory, accompanied by changes in burial customs, technology and mode of subsistence as well as the creation of new cross-continental networks of contact seen in the emergence of the pan-European Corded Ware Complex (CWC, ca. 2,900 to 2,300 calBCE) in Central and northeastern Europe. Studies of ancient genomes have shown that CWC were genetically closely related to the pastoralist Yamnaya Culture from the Pontic-Caspian steppe, bringing with them a genetic component that was not present in Europe previously. Genomes from the CWC of Central Germany suggest that this new genetic component replaced around 75% of the local Middle Neolithic genetic substrate. Presumably this ‘steppe’ genetic component spread in the subsequent millennia of the Final Neolithic and Bronze Age throughout Europe and can be seen in today’s European populations in a decreasing northeast to southwest gradient. Intriguingly, modern eastern Baltics carry the most WHG ancestry of all Europeans, supporting the theory of a remnant Mesolithic hunter-gatherer population in this region that left a lasting genetic impact on subsequent populations.

Conclusion With our analyses we support the pattern seen in the archeological record of continuity between the Mesolithic and Early Neolithic hunter-gatherer populations in the territory of modern Lithuania who appear genetically similar to Western hunter-gatherers. In contrast, contemporaneous hunter-gatherers from the more northern Latvia and Estonia were closer to Eastern hunter-gatherers. Networks of contact between the Baltic Sea and the river Volga could explain similarities seen in this region in pottery styles of hunter-gatherer groups although morphologically analogous ceramics could also have developed independently due to similar functionality. The situation appears differently in Scandinavia where a transition from foraging to agriculture in the Early Neolithic is carried by a demic diffusion from the south. Both the eastern Baltic and Scandinavia saw persistence of foraging throughout the Middle and Late Neolithic in populations that genetically largely descended from hunter-gatherer ancestors, as the resources of the Baltic Sea region, exploited through fishing and hunting, provided a beneficial environment for these groups and made it possible for them to maintain large population sizes without relying on crop cultivation. We see a population movement into the regions surrounding the Baltic Sea with the Corded Ware Complex in the Late Neolithic that introduced animal husbandry to the eastern Baltic regions but did not completely replace local foraging societies. The presence of ancestry from the Pontic Steppe among Baltic CWC individuals without the Anatolian farming component must be due to a direct migration of steppe pastoralists that did not pick up this ancestry in Central Europe. This could lend support to a linguistic model that sees a branching of Balto-Slavic from a Proto-Indo-European homeland in the west Eurasian steppe. As farming ancestry however is found in later eastern Baltic individuals it is likely that considerable individual mobility and a network of contact throughout the range of the CWC facilitated its spread eastward, possibly through exogamous marriage practices. Conversely, the appearance of mitochondrial haplogroup U4 in the Central European Late Neolithic after millennia of absence could indicate female gene-flow from the eastern Baltic region, where this haplogroup was present at high frequency.
(Source: “The Genetic History of Northern Europe”, by Alissa Mittnik et al.)
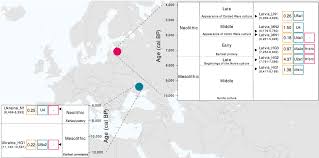
Summary The Neolithic transition was a dynamic time in European prehistory of cultural, social, and technological change. Although this period has been well explored in central Europe using ancient nuclear DNA, its genetic impact on northern and eastern parts of this continent has not been as extensively studied. To broaden our understanding of the Neolithic transition across Europe, we analyzed eight ancient genomes: six samples (four to ∼1- to 4-fold coverage) from a 3,500 year temporal transect (∼8,300–4,800 calibrated years before present) through the Baltic region dating from the Mesolithic to the Late Neolithic and two samples spanning the Mesolithic-Neolithic boundary from the Dnieper Rapids region of Ukraine. We find evidence that some hunter-gatherer ancestry persisted across the Neolithic transition in both regions. However, we also find signals consistent with influxes of non-local people, most likely from northern Eurasia and the Pontic Steppe. During the Late Neolithic, this Steppe-related impact coincides with the proposed emergence of Indo-European languages in the Baltic region. These influences are distinct from the early farmer admixture that transformed the genetic landscape of central Europe, suggesting that changes associated with the Neolithic package in the Baltic were not driven by the same Anatolian-sourced genetic exchange.
The Neolithic transition in the Baltic and Ukraine had a different tempo to that of central Europe, and it is unclear how this may have shaped the genetic composition of these regions.
The persistence of hunter-gatherer ancestry in the Baltic until at least the Middle Neolithic also provides a possible source for the resurgence of hunter-gatherer ancestry that is proposed to have occurred in central Europe from 7,000–5,000 cal BP.
Results suggest a degree of continuity across 4,000 years from the Mesolithic to the Neolithic period in the Dnieper Rapids.
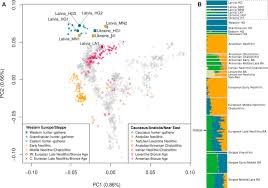
The ancient Latvian and Ukrainian samples fall close to modern Northern and Eastern European populations in PCA analysis, suggesting a degree of continuity in both regions since the Mesolithic period.
The Y chromosomes of two of our Latvian Mesolithic samples were assigned to haplogroup R1b (the maximum-likelihood sub-haplogroup is R1b1b), which is the most common haplogroup found in modern Western Europeans. This haplogroup has been found at low frequencies before the Late Neolithic in Western Europe but at higher frequencies in Russia and is suggested to have spread into Europe from the East after 5,000 cal BP. The presence of this haplogroup in Mesolithic Latvia points to a more westward ancestral range. We found that the three Mesolithic Latvian samples are predicted to have had the derived variant (rs12913832) of the HERC2 gene associated with blue eye color. Blue eye color is found at high frequencies in Northern Europe today, and these results suggest that this phenotype was already present in the Baltic by the Mesolithic period.
Discussion The Neolithic transitions in the Baltic and Dnieper Rapids region of Ukraine show very different archaeological and genetic dynamics to those observed in Central and Western Europe. Although in central Europe pottery and agriculture arrive as a package, in the Baltic and Dnieper Rapids the onset of the Neolithic is characterized by the appearance of ceramics, with a definitive shift to an agro-pastoralist economy only occurring during the Late Neolithic/Bronze Age.
We also find indications of genetic impact from exogenous populations during the Neolithic, most likely from northern Eurasia and the Pontic Steppe. These influences are distinct from the Anatolian-farmer-related gene flow found in central Europe during this period. It is interesting to note that even in outlying areas of Europe, such as Sweden and Ireland, an Anatolian-farmer-related genetic signature is present by the Middle to Late Neolithic period (∼5,300–4,700 cal BP). We conclude that the gradual appearance of features associated with the Neolithic package in the Baltic and Dnieper Rapids was not tied to the same major genetic changes as in other regions of Europe. The emergence of Neolithic features in the absence of immigration by Anatolian farmers highlights the roles of horizontal cultural transmission and potentially independent innovation during the Neolithic transition.
(Source: “The Neolithic Transition in the Baltic Was Not Driven by Admixture with Early European Farmers”, by Eppie R. Jones et al.)
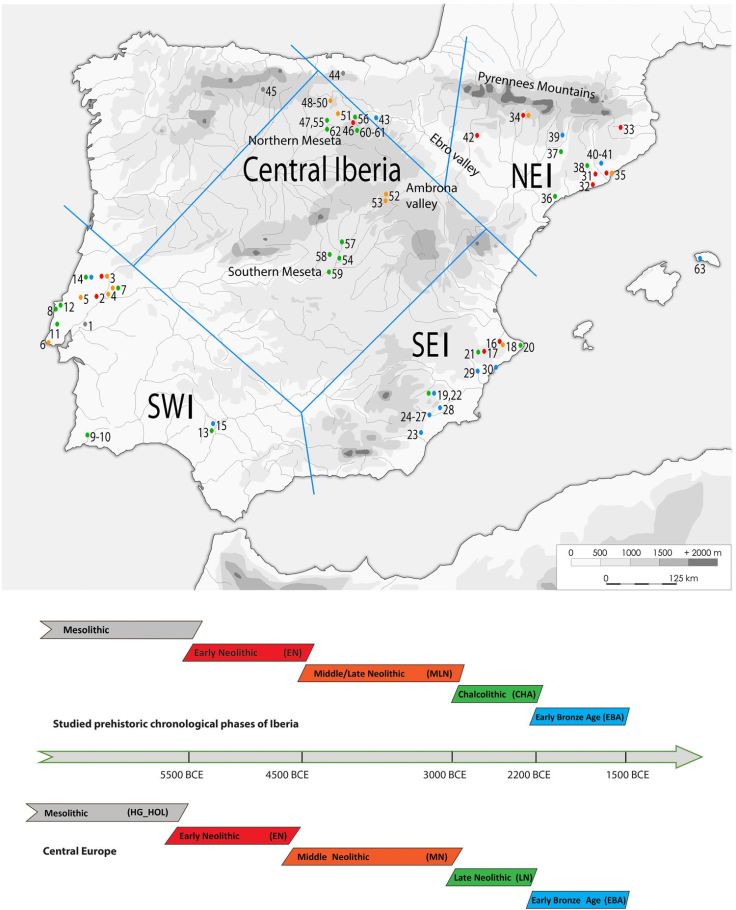
Abstract Agriculture first reached the Iberian Peninsula around 5700 BCE. However, little is known about the genetic structure and changes of prehistoric populations in different geographic areas of Iberia. In our study, we focused on the maternal genetic makeup of the Neolithic (~ 5500-3000 BCE), Chalcolithic (~ 3000-2200 BCE) and Early Bronze Age (~ 2200-1500 BCE). We report ancient mitochondrial DNA results of 213 individuals (151 HVS-I sequences) from the northeast, middle Ebro Valley, central, southeast and southwest regions and thus on the largest archaeogenetic dataset from the Peninsula to date. Similar to other parts of Europe, we observe a discontinuity between hunter-gatherers and the first farmers of the Neolithic, however the genetic contribution of hunter-gatherers is generally higher and varies regionally, being most pronounced in the inland middle Ebro Valley and in southwest Iberia. During the subsequent periods, we detect regional continuity of Early Neolithic lineages across Iberia, parallel to an increase of hunter-gatherer genetic ancestry. In contrast to ancient DNA findings from Central Europe, we do not observe a major turnover in the mtDNA record of the Iberian Late Chalcolithic and Early Bronze Age, suggesting that the population history of the Iberian Peninsula is distinct in character.

The artefactual remains, mainly ceramic, attest to the different origins and modes of Neolithisation on the Iberian Peninsula. On one hand stands a Mediterranean maritime colonization by Neolithic pioneers characterized by ceramic with clear parallels to the
Ligurian Impressa collections of Italian origin. Some Early Neolithic sites were also located in the hinterland, suggesting further routes of dissemination through the Pyrenees and/or along major rivers, such as the Ebro. On the other hand, North African influences and contacts are tangible in the southern Iberian Neolithic. All in all, Iberia appears a melting pot of influences and groups, combining Neolithic lifeways and with indigenous mechanisms of adaptation.
In the Early Neolithic, we can observe common features shared over large areas, but also some regionally restricted phenomena. In the whole territory, for example, there existed sophisticated systems of agriculture and livestock handling, with adaptable crops and seasonal strategies in flock management. The groups of the Franco-Iberian Cardial and the Epicardial pottery styles appeared at this time and recent studies have revealed mutual diachronic influences in the material culture and economies of these cultures.
From the late 5th millennium BCE onward, burial monuments appeared, and megalithic tombs became widespread. This phenomenon links Iberia with other parts of Europe, indicating long-distance networks of communication. Meanwhile, certain parts of northeast Iberia maintained individual inhumations in pits, mostly in small cemeteries. Besides megalithic tombs, ditched enclosures extending over more than 100 hectares started to dominate the landscapes of southern and central Iberia from 3300 to 3100 BCE, highlighting another widely spread European phenomenon. During the Iberian Chalcolithic period (3000-2200 BCE), fortified settlements with stone walls and semi-circular bastions appeared in the western and southern parts of the Peninsula, while elsewhere, open settlements were still extant. The diversity in settlement and burial types suggests the existence of social structures with different levels of complexity. At the same time, as exchange networks, circulated precious goods such as ivory from Africa and even Asia to Iberia.
Around 2200 BCE, the Chalcolithic settlement and funerary practices were suddenly discontinued, particularly in the western and southern part of Iberia, where most of the ditched and fortified settlements were abandoned and collective sites and megalithic tombs were replaced by individual burials. El Argar groups began to emerge in southeast Iberia, with large and massively fortified urban centers like La Bastida (Murcia), which managed to control a territory of over 35,000 km² during the following 650 years.
The northern part of Spain (Cantabrian fringe) attested to a rather complex Neolithic transition. Recent mtDNA and genome-wide analyses have put an emphasis on the genetic affinity and shared Near Eastern ancestry between the early Iberian farmers and the contemporary Central European Linearbandkeramik (LBK) population. The latest mtDNA and genomic studies have revealed increased subsequent admixture of hunter-gatherer elements during the local middle Neolithic (La Mina; Alto de Reinoso) and the Chalcolithic (El Portalon; El Mirador) again reminiscent of processes observed in Central Europe.
Despite numerous research projects being carried out over the past years, the Neolithic settlement history of Europe can still only be explained at a broad scale. Regional transects though time detailing the developments and the course of the Neolithic in central Germany and in the Carpathian Basin have mostly been examined by mitochondrial DNA (mtDNA) control region data. Our project completes the latter series by focusing on the archaeological models and hypotheses that have been put forward for the Iberian Peninsula, and where diachronic (i.e. ‘through time’) sampling of ancient DNA allows the detection of demographic changes and discontinuities between 5700 and 1500 BCE. Essential research questions focus on three levels: i) the individual sites, ii) the Iberian Peninsula as a scene of Neolithic transition and iii) a comparison with contemporaneous ancient and modern-day Europeans. A key question of our study of the mtDNA diversity on the Iberian Peninsula through time was to examine to what extent regional and supra-regional cultural groups could be recognized as genetically identifiable entities, as shown in other areas of Europe. A related question was whether the cultural breaks that can be seen, for instance, at the end of the Neolithic and Chalcolithic periods, were also accompanied by human population turnovers.

Conclusion The present study, based on 213 new and 182 published mtDNA data of prehistoric Iberian individuals, reveals a markedly different mode of interaction between local hunter-gatherers and incoming early farmers during the Early and Middle Neolithic of the Iberian Peninsula, as compared to Central Europe. As a characteristic of Iberian population dynamics, the proportion of autochthonous hunter-gatherer haplogroups was already high among the early Neolithic groups and increased even further in relation to the distance to the Mediterranean coast. In contrast, the early farmers in Central Europe showed comparatively little admixture of contemporaneous hunter-gatherer groups. Already during the first centuries of Neolithic transition in Iberia, we observe a mix of female DNA linages of different origins. Earlier hunter-gatherer haplogroups were found together with a variety of new lineages, which ultimately derive from Near Eastern farming groups. On the other hand, some early Neolithic sites in northeast Iberia, especially the early group from the cave site of Els Trocs in the central Pyrenees, seem to exhibit affinities to Central European LBK communities. This variety of contemporaneous haplogroups suggests that Early Neolithic migrations were not conducted via maritime routes across the Mediterranean only, but could also have included movement along inland routes, e. g. across the Pyrenees from the north. The diversity of female linages in the Iberian communities continued even during the Chalcolithic, when populations became more homogenous, indicating higher mobility and admixture across different geographic regions. Even though the sample size available for Early Bronze Age populations is still limited, especially with regards to El Argar groups, we observe no substantial changes to the mitochondrial DNA pool until 1500 BCE. The expansion of groups from the eastern steppe, which profoundly impacted Late Neolithic and EBA groups of Central and North Europe, cannot (yet) be seen in the contemporaneous population substrate of the Iberian Peninsula at the present level of genetic resolution. This highlights the distinct character of the Neolithic transition both in the Iberian Peninsula and elsewhere and emphasizes the need for further in depth archaeogenetic studies for reconstructing the close reciprocal relationship of genetic and cultural processes on the population level.
(Source: “The maternal genetic make-up of the Iberian Peninsula between the Neolithic and the Early Bronze Age”, by Anna Szécsényi-Nagy et al.)

Abstract One of the greatest transitions in the human story was the change from hunter-
gatherer to farmer. How farming traditions expanded from their birthplace in the Fertile Crescent has always been a matter of contention. Two models were proposed, one involving the movement of people and the other based on the transmission of ideas. Over the last decade, paleogenomics has been instrumental in settling long-disputed archaeological questions, including those surrounding the Neolithic revolution. Compared to the extensive genetic work done on Europe and the Near East, the Neolithic transition in North Africa, including the Maghreb, remains largely uncharacterized. Archaeological evidence suggests this process may have happened through an in situ
development from Epipaleolithic communities or by demic diffusion from the Eastern Mediterranean shores or Iberia. In fact, Neolithic pottery in North Africa strongly resembles that of European cultures like Cardial and Andalusian Early Neolithic, the southern-most early farmer culture from Iberia. Here, we present the first analysis of individuals’ genome sequences from early and late Neolithic sites in Morocco, as well as Andalusian Early Neolithic individuals. We show that Early Neolithic Moroccans are distinct from any other reported ancient individuals and possess an endemic element retained in present-day Maghrebi populations, indicating long-term genetic continuity in the region. Among ancient populations, early Neolithic Moroccans share affinities with
Levantine Natufian hunter-gatherers (~9,000 BCE) and Pre-Pottery Neolithic farmers (~6,500 BCE). Late Neolithic (~3,000 BCE) Moroccan remains, in comparison, share an Iberian component of a prominent European-wide demic expansion, supporting theories of trans-Gibraltar gene flow. Finally, the Andalusian Early Neolithic samples share the same genetic composition as the Cardial Mediterranean Neolithic culture that reached Iberia ~5,500 BCE. The cultural and genetic similarities of the Iberian Neolithic cultures with that of North African Neolithic sites further reinforce the model of an Iberian intrusion into the Maghreb.

Genetic analyses have revealed that the population history of modern North Africans is quite complex. Based on our aDNA analysis, we identify an Early Neolithic Moroccan component that is restricted to North Africa in present-day populations, which is the sole ancestry in IAM samples. We hypothesize that this component represents the autochthonous Maghrebi ancestry associated with Berber populations. This Maghrebi component was related to that of Epipaleolithic and Pre-Pottery Neolithic people from the Levant. By 3,000 BCE, a European Neolithic expansion brought Mediterranean-like ancestry to the Maghreb, most likely from Iberia. Our analyses demonstrate that at least some of the European ancestry observed today in North Africa is related to prehistoric migrations, and local Berber populations were already admixed with Europeans before the Roman conquest.

(Source: “Neolithization of North Africa involved the migration of people from both the Levant and Europe”, by Rosa Fregel et al.)

NovoScriptorium: What we’ve learned from the study of the above sources is that we hardly know much about human evolution in terms of genetics. And we mean the actual truth, the undoubted fact(s) that we all must agree on. We’ve noticed quite a few contradictions between researches, many assumptions, however rational they may be, and, the worse of all, ‘dogmatic’ thinking as if some ‘assumptions’ are the ‘holy cows’ of Genetics and Archeology. The vast majority of the results are based on DNA extracted from a single or a couple of individuals. Even if the number of individuals increases, how do we know that he/she/they weren’t ‘isolated’ cases? How do we know that the DNA extracted is actually ‘readable’ and not distorted by other parameters? How do we know that we ‘read’ the DNA properly? Reading the DNA is probably the most difficult task for us humans, as the combinations and variations are virtually endless. How can we claim ‘genetic replacements’ while Archeological data shows cultural continuity in the same area of examination? Why are specific regions of Europe, in terms of their prehistory, provokingly understudied? How can we reach ‘general -core- conclusions’ while crucial areas are not yet studied properly? And the questions could keep coming for ever. With this we are not aiming to reject the work of excellent and respected scientists; on the contrary, we want to emphasize on the need to keep Science…scientific.
Research-Selection-Comments for NovoScriptorium: Maximus E. Niles

Reblogged this on Muunyayo .
LikeLike
Incredible work and presentation!
LikeLiked by 1 person